CHICAGO—The lymphatic system took the spotlight in the ACR Convergence 2025 session, Flushing the Joint: Using Lymphatic Drainage to Control Synovial Inflammation, in a basic science talk exploring the role of lymphatics in inflammatory arthritis pathogenesis. The session explored the structure and function of the synovial lymphatic network through a series of elegant experiments, revealing its critical role in modulating synovial inflammation. The talk was delivered by osteoimmunologist Edward Schwarz, PhD, the Burton Professor of Orthopedics and director of the Center for Musculoskeletal Research, University of Rochester, New York.
Introduction to the Lymphatic System
Dr. Schwarz opened the session with a thought-provoking question: If rheumatoid arthritis (RA) is a systemic disease, why is the second metacarpophalangeal (MCP) joint particularly prone to inflammation? As a potential explanation, he projected striking images of a hand injected with fluorescent dye to illustrate significant lymphatic drainage through the second MCP, more so than through the neighboring, smaller MCPs.1
The lymphatic system begins distally as a simple tube of capillaries, which drain interstitial fluid and inflammatory cells. These capillary tubes converge into afferent collecting lymphatic vessels, which are surrounded by lymphatic muscle cells capable of contraction. These collecting vessels contract to pump fluid to draining lymph nodes, which subsequently return fluid to the venous system.
To illustrate these lymphatic vascular contractions, he displayed images from a mouse model. Infrared indocyanine green (first used in oncologic surgery to identify sentinel lymph nodes) was injected into the footpad of a mouse, and lymphatic vessels were seen actively contracting and draining into the more proximal popliteal lymph node.
In acute arthritis mouse models, the contracture frequency of these lymphatic vessels is dramatically increased, while in chronic models, the contractions are lost. Dr. Schwarz encouraged attendees to imagine an acutely inflamed ankle due to rheumatoid arthritis: Considering the muscle cell-laden lymphatic vessels reside more proximally up the limb, how do the collecting lymphatic vessels know synovial inflammation is present distally and initiate contractions?
Expanding & Collapsed Lymphatic Phases
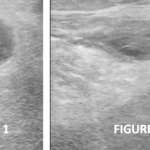
In 2007, Proulx et al. demonstrated that in a mouse model of ankle synovitis, the popliteal lymph node was 10–20 times its normal size. They went on to describe the “expanding” and “collapsed” phases of lymph nodes in inflammatory arthritis.2
The expanding phase occurs prior to the onset of distal synovitis, in which there is a large, contrast enhanced, draining lymph node. Eventually, the lymph node will collapse and no longer take up intravenous contrast, which occurs concomitantly with a flare of distal synovitis.3 This expanding and collapsed mechanism also correlates with the velocity of macrophages traveling in the popliteal lymphatic vessels.4