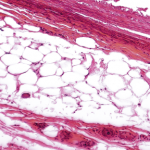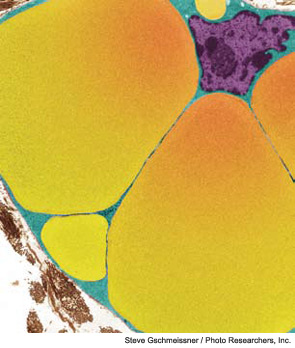
Colored transmission electron micrograph of lipid droplets (yellow) in a developing adipocyte.
A dipose tissue has longed been viewed as a harmless tissue in the pathogenesis of chronic inflammatory connective tissue and joint diseases, with fat providing the soft surroundings for damage inflicted by other mechanisms. This dogma, based on the protective role of leptin in metabolic diseases, has long been popular, although it may have hindered thorough basic science research in this field. However, three discoveries have completely changed this point of view and will be the subject of this article.1,2
These advances can be summarized briefly as follows: